 Makroevlusjon-filosofi og empiri
Makroevlusjon-filosofi og empiri
John C. Lennox viser i sin bok 'Gods undertaker' at langt fra å være utledet fra vitenskapelige forsøk og empiri, så er teorien om felles avstamning bygd på naturalistisk filosofi: Det har vist seg tilfelle i årtusener at evolusjon, av materialistisk-naturaltistisk nødvendighet har presset fram forklaringer. Den greske materialistiske filosof Epikur, anvendte nettopp denne filosofien til å anvende en evolusjonistteori ut fra Demokrits atomteori. Den romerske dikter Lucretius ga det mest kraftfulle uttrykk for epikureisk teori i diktet 'Om Universets natur' fra ca år 50 f.Kr. Han er blitt kalt 'den første darwinist' og hans filosofi ble gjenopplivet med begeistring i renessansen. Noen mener han bør regnes som stamfar til moderne naturalistisk filosofi. Biologisk makroevolusjon står i tillegg i så tett forhold til naturaltistisk filosofi at det kan utledes direkte av den. Lucretius gamle argument i den retning viser det. Det er vanskelig å tenke seg en annen vitenskapelig teori som befinner seg i samme situasjon. Det ville være vanskelig å utlede Newtons teori om gravitasjon eller Einsteins relativitetsteori ut fra et filosofisk prinsipp eller verdensanskuelse. (Mer her.)
Hvor godt er det vitenskapelige grunnlaget for å hevde makrovolusjon? Evolusjon i forbindelse med makroevolusjon henviser til en form for nyskapelse, noe med framkomst av nye organer, kroppsstrukturer, noe med kvantitativt nytt genetisk materiale etc. Makroevolusjonær forandring involverer altså en betydelig forøkning av kompleksitet. I tillegg til organer, skal proteiner for disse dannes -hvilket igjen kan kreve dobbeltmutasjoner, om det skal forklares ved darwinistiske mekanismer. Tesen om glidende overganger forutsetter at makroevolusjon forklares ved de samme prosesser som driver mikroevolusjon, bare det er nok tid til rådighet. Paul Wesson har framsatt en erklæring som eksemplifiserer viktigheten av en sondring mellom mikro-og makro -evolusjon: 'Store evolusjonære nyskapelser har vi ikke særlig godt grep om. Vi har aldri observerte en eneste av dem, og vi har ingen anelse om hvorvidt noen overhodet er i stand til å utfolde seg. Det finnes heller ingen fossile vitnesbyrd om det. Til sammenligning er mikroevolusjonære variasjoner, forårsaket av mutasjoner og naturlig utvalg observert til overmål.
Har eksperimenter bekreftet makroevolusjon?
Om en går inn på hva eksperimenter viser, kan vi se litt på når slike endringer knyttet til kroppsbygning må inntreffe: For å danne betydelige endringer i kroppsbygningen til dyr, er det viktig hvor tidlig i embryo-stadiet mutasjoner inntrer. Det er jo fordi sent i utviklingen, så er allerede hovedtrekkene i kroppsbygningen fastlagt. Sentvirkende mutasjoner kan derfor ikke framkalle store endringer. Således er det bare mutasjoner framkalt tidlig i embryo-stadiet som har noen sjanse til å avstedkomme storskala makroevolusjonær forandring. Dette bekreftes av flere evolusjonær-biologer (G. Miklos, K.Thompson ) Likevel i eksperimenter fra tidlige på 1900-tallet (T.H.Morgan..) , inntil i dag , så har erfaringen ensidig vært ødeleggende for organismen utsatt for mutasjon. Slike mutasjoner er enten dødelige eller resulterer i organismer som ikke kan leve i vill tilstand (neo-darwinist pioner R.A.Fisher) En endring tidlige i embryo-stadiet, ville kreve en rekke andre koordinerte forandringer i atskilte, men funksjonelt interrelaterte utviklingsprosesser. Det er den tette integreringen av funksjoner gjør at tidlige mutasjoner fører til død i embryo-stadiet. (Se evt. mer her.)
 Dannelse av 3-dimensjonale protein-folder
Dannelse av 3-dimensjonale protein-folder
Douglas Axe ønsket å benytte sin innsikt som ingeniør til å fokusere på ny genetisk informasjon nødvendig for å danne nye protein-folder, som kritiske tester på neo-darwinistiske mekanismer. Proteiner innbefatter minst tre ulike struktur-nivåer(4): Den spesifikke kjeden av aminosyrer i protein, eller polypeptid-kjeden er den primære strukturen. Den andre strukturen utgjøres av gjentagne alfa-spiraler eller beta-strands (eng). Den tredje tilsvarer en 3-dimensjonal protein-folding. Axe visste at i det nye livsformer oppsto i livets historie, slik som i den kambriske eksplosjon, så måtte mange nye proteiner ha oppstått. Nye dyrearter har nye organer og nye celle-typer, og nye celle-typer krever nye proteiner for å betjene dem. I noen tilfeller kunne nye funksjonelle proteiner, fungere med samme tre-dimensjonale folding som tidligere proteiner. Men i de fleste tilfeller så krevde nye fungerende proteiner, ny folding for å utøve sine funksjoner. Utbrudd av nye livsformer {f.eks. i forbindelse med økt oksygenandel} måtte dermed også ha resultert i nye former også av protein-folding.
Genetikeren og evolusjonær biolog, Susumo Ohno, bemerket at den kambriske eksplosjon krevde komplekse nye proteiner, f.eks. lysyl oxidase, for å støtte deres omfattende kropps-struktur. Da disse proteinene inntraff i kambriske dyr, sammentraff de sannsynligvis med nye typer av protein-folding. Axe ble dermed overbevist om at å forklare denne type innovasjoner, krevde en mekanisme som i det minste kunne produsere ny protein-folding. Som ingeniør så han at nye kroppsformer krevde innovasjon i form og struktur. Som vitenskapsmann i forhold til proteiner, så han at ny protein-folding kunne bli sett som den minste strukturelle innovasjon i livets historie. Derfor måtte mutasjoner generere nye protein-foldinger, for at naturlig seleksjon kunne ha anledning til å bevare og akkumulere strukturelle innovasjoner. Axe anså dermed at evnen til å produsere nye protein-foldinger representerte noe' en ikke kunne klare seg foruten', ett 'sine qua non' i makroevolusjonær innovasjon. Axe innså at hvorvidt mutasjoner kunne danne nye protein-foldinger, avhang av hvor sjeldne funksjonelle gener og proteiner var i det mulige 'rommet' av mulige sekvens-følger, og hvorvidt tilfeldige genetiske mutasjoner ville kunne gjennomsøke det relevante sekvens-rommet i løpet av evolusjonær tid.
I artikkelen til Sauer et al, estimerte de forholdet av funksjonelle proteiner å være ekstremt lavt (1 til 10 opphøyd i 63). Axe bemerket at forfaterne valgte ikke å understreke sjeldenheten, men heller variasjonen av aminosyre-substitutter som aktuelle proteiner kunne fungere under. I artikkelen fremholdt Sauer et al den daværende populære hypotesen at det var aminosyrene i det indre av et foldet protein som var viktigst, mens de ytre ikke betød på langt nær så mye. (5) Axe utførte et eksperiment for å teste dette, og ble overrasket over resultatet. Da Axe erstattet hele det innholdet i et lite, internt enzym med tilfeldig sammensatte aminosyrer, så utførte likevel en femdel sin opprinnelig funksjon. Dette antydet at proteiner kanskje var mindre følsomme for funksjonelle tap, som følge av sekvensendringer, enn Axe hadde trodd.
I neste omgang fokuserte han på det ytre av proteiner, i det han tilfeldig omgjorde to proteiners ytre, på samme vis som han hadde endret det indre til ett av dem Denne gangen feilet resultatet å produsere noen funksjonelle varianter i det hele. I det Axe innså at dette motsa hva Sauer et al hadde kommet fram til, besluttet Axe å gjøre mye mindre endringer i neste forsøk. Han erstattet hver ytre aminosyre bare med det mest like alternativet. Likevel mistet begge proteinene han studerte all funksjonalitet, innen han hadde erstattet en femdel av deres ytre fremtoning. Han konkluderte ut fra dette, at de ytre delene av proteiner var mye mer utsatt for funksjonelt tap, enn hva som hadde vært formodet. Axe disponerte på laben mer sensitivt verktøy som tillot å fastslå at selv om enkelt-mutasjoner tillot proteiner å beholde noe funksjonalitet, så minket eller ødela de likevel funksjonaliteten ofte nok til å sikre at ville elimineres ved den rensende effekten til naturlig seleksjon. Axe lærte også at enhver enkelt mutasjon som feilet i testen hans, på egen hånd ødela proteinets funksjonalitet. Han fant ut at 5 % av slike enkeltendringer ødela protein-funksjonaliteten. Som konklusjon viste han at proteiner, til tross for noe tillatt variasjon, er høyst spesifike m.h.t. til sin biologiske funksjon. (Les evt. mer her.)
Hva med fossiler?
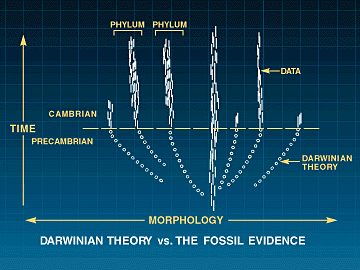 Stephen Jay Gould har uttalt: 'At vi har funnet så ekstremt få overgangsformer i den fossile lagserie, er fortsatt paleontologenes forretningshemmelighet. Hans fagkollega i 80-årene, Niles Elderidge tilføyer: 'Når vi endelig ser en evolusjonær nyhet duke opp, skjer det normalt med et brak. Ofte er det uten noen håndfast form for bevis på at fossilene ikke har utviklet seg andre steder. Han kommer med en bemerkelsesverdig innrømmelse: 'Faktisk er det slik at vi paleontologer har sagt at livet historie understøtter (historien om gradvis adaptive tilpasninger), mens vi utmerket har vært klar over at den ikke gjør det.
Stephen Jay Gould har uttalt: 'At vi har funnet så ekstremt få overgangsformer i den fossile lagserie, er fortsatt paleontologenes forretningshemmelighet. Hans fagkollega i 80-årene, Niles Elderidge tilføyer: 'Når vi endelig ser en evolusjonær nyhet duke opp, skjer det normalt med et brak. Ofte er det uten noen håndfast form for bevis på at fossilene ikke har utviklet seg andre steder. Han kommer med en bemerkelsesverdig innrømmelse: 'Faktisk er det slik at vi paleontologer har sagt at livet historie understøtter (historien om gradvis adaptive tilpasninger), mens vi utmerket har vært klar over at den ikke gjør det.
Hva er det da fossilhistorien avslører? Gould har skrevet: De fleste arters historie omfattes av to forhold som er i loddrett motstrid med idéen at de skal ha utviklet seg gradvis.
1. Stillstand. De fleste arter utviser ingen retningsbestemte endringer i løpet av deres livstid her på jorda. De dukker opp i den fossile lagserien stort sett med samme utssende som når de forsvinner igjen. Den morfologiske forandring er som regel begrenset og uten retning.
2. Plutselig tilsynekomst. Ikke på et eneste sted oppstår en art gradvis ved en stabil forandring av sine forfedre. De framtrer på scenen uten foregående tillp og 'fullt ferdige'.
Måten Gould og Eldredge ser den fosile lagserie, med korte perioder med hurtige forandringer og så lange perioder med stillstand, har fått dem til å fremsette en teori 'avbrutt likevekt' (punktuated equilibrium'). Som et eks. peker Gould i sin bestselger 'Wonderful Life' på hvordan alle de store taksonomiske rekker vi har i dag, samt noen utdødde, plutselig dukket opp i den såkalte Kambriske eksplosjon. Dette øker vanskelighetene for dem som vil argumentere for at mikroevolusjonære prosesser utgjør en tilstrekkelig motor for makroevolusjon.
Til tider har det vært strid mellom gradualismen i neo-darwinismen og den 'avbrutte likevekts' teorien. Gradualistene holder på at mikroevolusjonen over et visst tidsrom blir til makroevolusjon. Mens Eldredge holder dem for å være ganske blanke, hva paleontolgi gjelder. Hans argument er at selv om gradualistene gjør meget ut av å få fram hvordan genetisk informasjon er endret i årenes løp, så ender det med at slår seg til ro med at evolusjonshistorien er et resultat av det naturlige utvalgs arbeid med allerede tilgjengelig genetisk variasjon.
Man kan ikke ta utgangspunkt i hva vi kan obserere i nåtiden, og så kjøre baklengs gjennom geologisk tid. "En slik simpel ekstrapolering duger simpelten ikke", sier Eldredge. Denne kjennelse fra sakkunnskapen, som er i så påfallende motstrid mot gjengse forestillinger om fossiller, støttes av Colin Patterson (FRS): "Jeg vil ikke stikke noe under stolen: Det finnes ikke her ett eneste sådant fossil (som brukes som 'stamfar'eller overgangsform), hvormed man vil kunne lage et vanntett argument". Det er interessant i og med den såkalte urfuglen (Archaeopteryx) befant seg i han varetekt på Natual History Museum. Her skal også tas med Patersons begrunnelse, som jeg nå er gjort oppmerksom på: "“Årsaken er at utsagn om forfedre og nedstigning ikke er aktuelt i fossilregistreringen. Er Archeopteryx forfedre for alle fugler? Kanskje ja, kanskje nei: det er ingen måte å svare på spørsmålet på." (1) {Dette skyldes bl.a. at fossiler ikke kommer med ID-papirer, og det ikke er mulig i utgangspunktet og si at det ene fossilet stammer direkte fra et annet -oversetters kommentar.} Evt. mer her.
Som kristen har en i det minste ett valg for hvordan en vil se for seg menneskelig avstamning, men at det kan komme noe godt ut av naturlige, ikke-styrte prosesser, er det opp til evolusjonsteoriens meningsbærere å vise.
Samlet og redigert av Asbjørn E. Lund